Previous: Transport across the tropopause by Rossby wave breaking Next: Effective diffusivity Up: Ext.Abst.
4. Stretching rates
One possible measure of the mixing propertes of a given atmospheric flow is the stretching rates of materially advected contours, related to the Liapunov exponents of the flow (eg. Ottino 1989) and essentially the rate at which fine scales are produced on which molecular diffusivity may operate. On isentropic surfaces a suitable choice of material contour are isolines of potential vorticity. Further, considering the tropopause as a transport barrier suggests the possibility of identifying it with a PV contour that exhibits a minimum stretching rate. Bithel & Gray (1997) considered stretching rates for a six day period in October 1990, using contours initialized with a range of PV values, but found no single value of PV that yielded a minimum stretching rate, and that could therefore be identified with the tropopause.
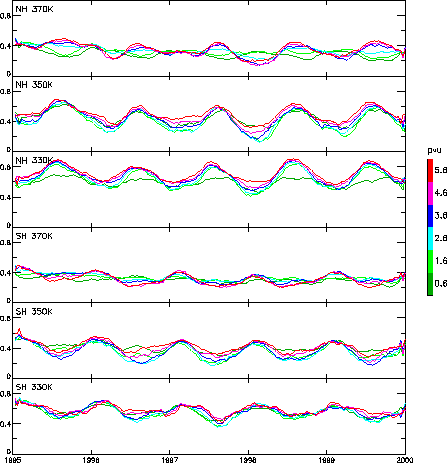
Here, we reexamine the contour stretching rates near the tropopause, but this time focus on broader scale patterns and their seasonal and interannual variability. We performed a series of integrations (with no surgery) over a 5 year period from 1995 to 1999, with three integrations per month, and on each of the isentropic surfaces 330K, 350K, 370K. The contours were initialized with PV from ECMWF analyses taking values of (±) 0.6, 1.6, 2.6, 3.6, 4.6, 5.6 pvu. Each integration lasted 6 days, which was found to be sufficient to define a stretching rate. Figure 2 shows the stretching rates of each simulation, plotted as a function of initial time, after smoothing with a boxcar of width 70 days (or 7 integrations). Unsmoothed results (not shown) revealed considerable short term (10 day) variability, consistent with the time-scale of baroclinic lifecycles or Rossby wave breaking events.
Several features of Figure 2 are worth noting. First there is a clear annual signal at all levels, with stronger mixing in the summer and weaker mixing in the winter, in accordance with our current understanding of the seasonality of the tropopause (eg. Appenzeller et al., 1996), as well as with more specific recent studies of, eg. effective diffusivity (Haynes & Shuckburgh, 2000). Although this annual signal is in general weaker at 370K, it is still visible there, especially in the higher PV contours of ((±)) 3.6, 4.6, 5.6 pvu. Note also that the 0.6 and 1.6 pvu contours at 370K in the NH show an annual cycle that is in antiphase with the higher PV contours, ie with mixing strongest in winter, weakest in summer.
Another important feature of Figure 2 is the difference in stretching rates between different PV contours at a particular time. For example, at 350K in both hemispheres there is a wider range of stretching rates in the winter than in the summer. Thus, considering the tropopause as a dynamical barrier to mixing, the stretching rates in the winter portions of Figure 2b,e, suggest identifying the tropopause with 1.6, 2.6 or 3.6 pvu contours, eg 2.6-3.6 in the SH 1995 and 1996 winters, or 1.6-2.6 in the NH winters. Similarly such arguments suggest a strong seasonality of the tropopause at 370K, varying over the full range of PV contours used here, from 5.6 pvu in the winter to 0.6 pvu in the summer.
Considering interhemispheric differences in general, we note that that stretching rates are higher in the NH than in the SH, most notably at 330K and 350K. This is consistent with the generally more disturbed troposphere in the NH, associated with the stronger topography and land-sea temperature contrasts.
Finally, although the time-series is too short to infer any long term trends, Figure 2 gives some suggestions of the interannual variability. For example, the NH winter at the beginning of 1998, following a strong El Nino event, is markedly more stable at all levels than the surrounding winters, while both the NH and SH winters of 1995 is less stable, most notably at 350K and 370K. Further calculations extending the time-series backwards are necessary to identify any possible relation between the stability of the tropopause region and, eg., the phase of ENSO.
Figure 2: Contour lengthening rates the 330K, 350K and 370K isentropic surfaces in each hemisphere, in units of 1/day.
Previous: Transport across the tropopause by Rossby wave breaking Next: Effective diffusivity Up: Ext.Abst.